Голосеменные
Спешу обрадовать, мы добрались до изучения семенных растений! К ним относятся голосеменные и покрытосеменные (цветковые). До этого размножение шло только с помощью спор: у мхов, папоротников, хвощей и плаунов — высших споровых растений. Настало время открыть новую интересную главу этой книги, посвященную растениям, которые размножаются с помощью удивительного изобретения природы — семени.
Голосеменные — распространенная древняя группа растений, включающая небольшое число видов. Главной особенностью данной группы являются «голо» (то есть открыто) лежащие семяпочки и, в дальнейшем, развивающиеся из них семена. Иными словами, у голосеменных растений отсутствуют замкнутые вместилища для семян.

На примере типичного представителя — сосны обыкновенной, относящейся к классу хвойных, поговорим о характерных чертах данного класса и голосеменных растений в целом.
Общие признаки
- Деревья и кустарники
Все голосеменные представлены древесными формами: деревьями и кустарниками. Травы отсутствуют.
Хвоинки (хвоя) — игольчатые видоизменения листьев. Сохраняются долгие годы, у некоторых сосен до 45 лет. Хвоя лиственниц опадает ежегодно.

Древесина голосеменных обладает большим запасом механической прочности. Это связано с ее особенностями: она состоит из трахеид с окаймленными порами, паренхима развита слабо. Либриформ (древесные волокна) и настоящие сосуды отсутствуют (исключение — гнетовые, имеют сосуды). Клетки-спутницы во флоэме также отсутствуют.
В древесине и коре имеются каналы, заполненные смолой. Однако, есть исключения — у гинкго смола не образуется вовсе.
Несколько веков назад в России целенаправленно создавались и охранялись, так называемые, корабельные рощи. Это, прежде всего, требовалось для флота, так как мачты кораблей изготавливали из сосен, отвечающих всем требованиям — корабельных (гладкий, твердый и прочный прямой ствол с минимальным количеством сучков и смолы).

Семяпочки и развивающиеся из них семена лежат «голо», открыто, для них нет закрытых вместилищ, отсутствует завязь. В сравнении с высшими споровыми растениями, размножение семенами ставит голосеменных на более высокий уровень организации.
Голосеменным растениям для размножения не требуется вода, опыление у них происходит с помощью ветра. Этот процесс перестал быть зависимым от капельно-жидкой среды, как было у мхов и у папоротников. Благодаря этому голосеменные получили большое преимущество и смогли расселиться по всей Земле, в том числе в засушливых районах. Они господствовали в юрском периоде, когда климат стал более сухим и жарким.

Обитают голосеменные в местах с холодным климатом и достаточным количеством влаги. Имеются виды, обитающие в жарких странах: растение вельвичия удивительная обитает в пустынях южной Африки.
Строение и жизненный цикл
Жизненный цикл голосеменных состоит из чередования бесполого поколения — спорофита (диплоиден, 2n), и полового поколения — гаметофита (гаплоиден, n). Господствует (доминирует) в цикле спорофит (2n) — это взрослое растение сосны.
Голосеменные относятся к разноспоровым, как и все семенные растения. Они образуют разные споры: крупные женские (мегаспоры) и мелкие мужские (микроспоры). Образуются они в спорангиях, расположенных на спорофиллах, которые собраны в стробилы (шишки) — от лат. strobilus — сосновая шишка.
Мужские шишки (стробилы)
К концу весны у основания молодых побегов образуются мужские шишки (стробилы) — мелкие, собранные в тесные группы, желтого цвета. Чешуи мужских шишек представляют собой микроспорофиллы. Микроспорофиллы — гомологи тычинок, которые крепятся к оси каждой шишки спирально, с нижней стороны, и имеют два пыльцевых мешка — микроспорангия.
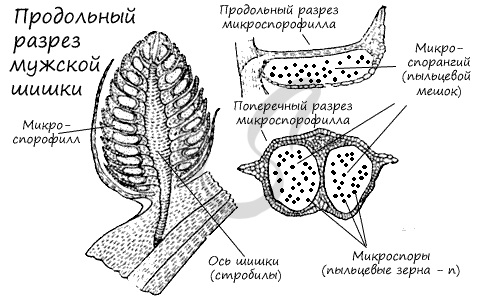
Образование мужского гаметофита
Из материнских клеток (2n) в микроспорангии путем мейоза образуются 4 микроспоры (n). Строение микроспоры следующее: она покрыта экзиной (от гр.exo снаружи, вне) — наружная оболочка, изнутри интиной (от лат. intus внутри) — внутренней оболочкой. В составе микроспоры имеются также два воздухоносных мешка, образованных в результате отслоения экзины от интины и возникновения полости между ними.
Микроспора делится, не покидая спорангия, преобразуется в заросток. При делении из ядра микроспоры образуются две клетки. Одна из них превращается в две заростковые клетки (протоллиальные — от греч. проталлиум — заросток) — быстро отмирают и исчезают. Их функция до конца не изучена.
Из другой клетки в ходе митоза также образуются две: антеридиальная, из которой развиваются мужские половые клетки — спермии (неподвижные, без жгутиков в отличие от сперматозоидов), и более крупная вегетативная клетка, из которой в дальнейшем формируется пыльцевая трубка.

Мужской гаметофит сильно упрощен, антеридии отсутствуют. Формируется он прямо внутри микроспоры, которая в итоге превращается в пыльцевое зерно. Совокупность пыльцевых зерен называется пыльца.
При вскрытии (нарушении целостности) микроспорангия, или пыльцевого мешка, пыльца высыпается во внешнюю среду и достигает женской шишки, где, в результате опыления, внутри семязачатка происходит дальнейшее развитие мужского гаметофита.
Образование женского гаметофита
На тех же самых соснах, где расположены мужские шишки, лежат и женские. Весной на верхушке молодого побега появляются мелкие (около 5 мм) красноватые шишки — это женские шишки (стробилы). Состоят они из оси (стержня) , на котором располагаются две чешуи: кроющая и семенная. На верхней стороне у основания семенной чешуи лежат два семязачатка.
Кроющая чешуя представляет собой видоизмененный лист, в его пазухе находится семенная чешуя. Семенная чешуя — видоизмененный боковой побег.
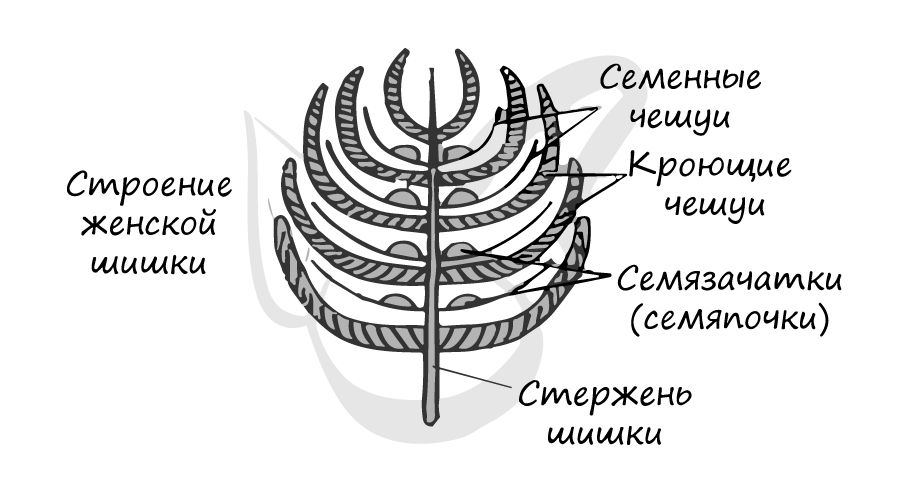
Женские шишки (стробилы)
Именно открыто расположенные семязачатки (семяпочки) служат причиной, по которой этот отдел растений называется — голосеменные.
В женских шишках, в отличие от мужских, каждая чешуя гомологична целой мужской шишке (стробилу). То есть одна чешуя — целой мужской шишке, а не отдельным ее микроспорофиллам (чешуям)!
Молодой семязачаток состоит из нуцеллуса, интегумента и фуникулуса. Нуцеллус (от лат. nucella — орешек) — центральная часть семяпочки, соответствующая мегаспорангию. Интегумент (от лат. integumentum покрывало) — покров семяпочки, вырастающий из ее центральной части — нуцеллуса. В зрелом семени интегумент преобразуется в семенную кожуру. Фуникулус (от лат. funiculus канатик, верёвка) или семяножка — часть семязачатка, соединяющая его с мегаспорофиллом (семенным чешуями).
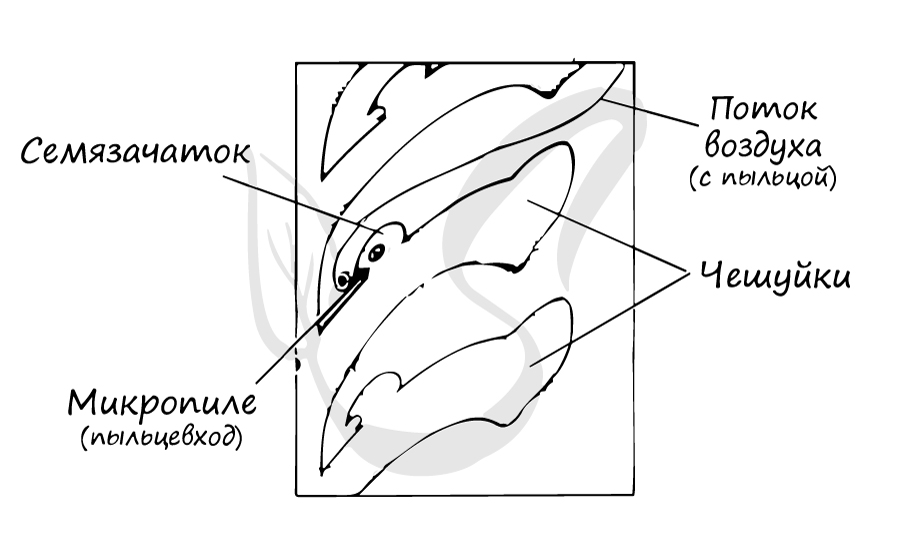
На интегументе около вершины располагается микропиле (пыльцевход) — через него после опыления пыльцевая трубка проникает в нуцеллус. Между интегументом и нуцеллусом имеется густая жидкость, выступающая из микропиле. Подсыхая, она втягивается внутрь семязачатка и затягивает вместе с собой пыльцу, осевшую на ней.
Образование женского гаметофита
В средней части обособляется спорогенная клетка (2n) (археспориальная — от греч. arche начало и sporá семя). В результате ее митотического деления образуются материнские клетки спор — спороциты (2n), однако и сама археспориальная клетка может выступать в роле спороцита, минуя стадию митоза. Спороциты (2n) делятся мейозом на четыре гаплоидные (n) мегаспоры.
Три мегаспоры отмирают, остается одна, которая многократно делится митозом и формирует эндосперм — запасное питательное вещество. Обратите на этот факт особое внимание: у голосеменных эндосперм гаплоидный (n) и образуется до оплодотворения. Такой эндосперм называется — первичный, он соответствует женскому гаметофиту.
Как и мужской, женский гаметофит весьма упрощен и заключен внутри мегаспоры. На верхушке женского гаметофита (мегагаметофита) образуется архегоний с яйцеклеткой (n). У гнетовых архегонии отсутствуют.

Жизненный цикл
На спорофите (2n) в микроспорангиях из материнских клеток (2n) путем мейоза образуются микроспоры (n). Из микроспоры формируется пыльцевое зерно. Пыльца (пыльцевые зерна (n)) с помощью ветра попадает в женские шишки, где улавливается густой жидкостью между интегументом и нуцеллусом, выступающей из микропиле. Жидкость засасывает пыльцу внутрь семязачатка на нуцеллус (в пыльцевую камеру). После того, как опыление произошло, микропиле зарастает. Чешуи шишки смыкаются и склеиваются смолой.
Семязачатки в этот момент еще не готовы к оплодотворению, так что от момента опыления до оплодотворения проходит около 13 месяцев. За это время в семязачатке формируется эндосперм, женская шишка увеличивается до 3-4 см и приобретает зеленую окраску.
Оказавшись на мегаспорангии, наружная оболочка пыльцевого зерна (экзина) разрывается, из вегетативной клетки в направлении архегония начинает расти пыльцевая трубка. Антеридиальная клетка делится на генеративную (спермагенную) и клетку-ножку антеридия (функция последней до сих пор не изучена). Спермагенная клетка попадает в пыльцевую трубку, а из нее — в архегоний.
Непосредственно перед оплодотворением спермагенная клетка делится на два спермия (n), один из которых отмирает, а другой сливается с яйцеклеткой (n). Образуется зигота (2n), из которой формируется и растет зародыш благодаря эндосперму — запасу питательных веществ.
Окончательно созревают семена к осени на второй год после опыления, к этому моменту женские шишки увеличиваются в размерах до 6 см. Зеленая окраска меняется на серую, чешуйки расходятся, и семена, образовавшиеся из семязачатков, высыпаются. Из семени прорастает взрослое растение — спорофит (2n). Цикл замыкается.
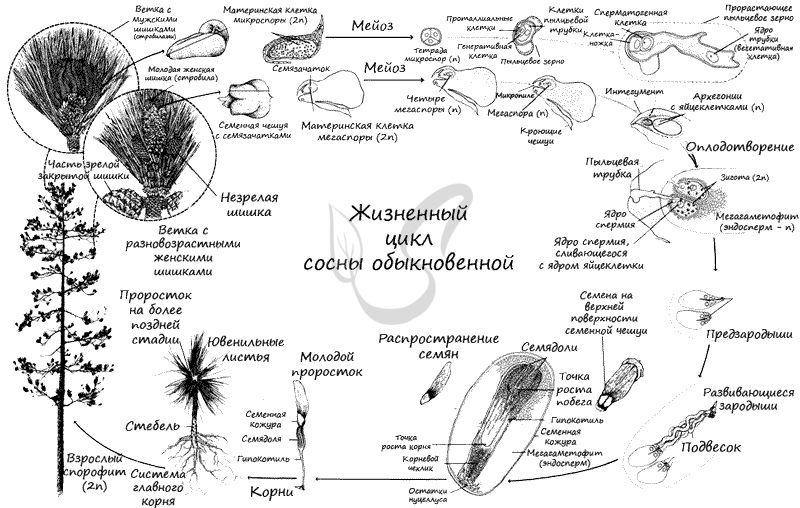
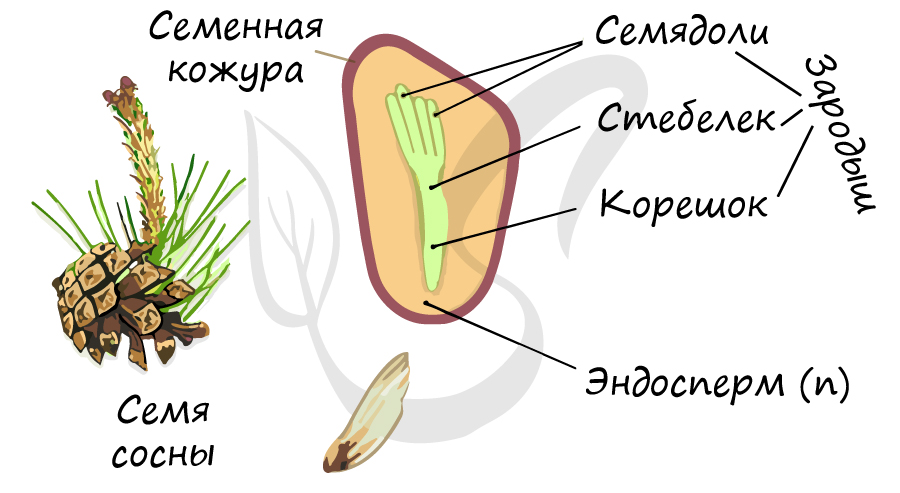
Строение семени
Семенная кожура, защищающая семя от пересыхания и неблагоприятных факторов внешней среды, образована разросшимся интегументом.
Зародыш (2n) формируется в результате митотического деления образовавшейся зиготы. Состоит из зародышевого корешка, стебелька и почечки.
Число семядолей у голосеменных различается — от 2 до 15. Семядоли имеют доступ к запасным питательным вещества (эндосперму).
Запасные питательные вещества накапливаются в эндосперме (n). Особенностью в строении семени голосеменных, по сравнению с семенем покрытосеменных (цветковых) является наличие гаплоидного эндосперма (n). Не забывайте, что эндосперм у голосеменных это производное мегагаметофита (n), исходя из этого становится понятно, почему ткань гаплоидна. У цветковых, в отличие от голосеменных, эндосперм триплоиден (3n).
Фитонциды
Фитонциды (от греч. phyton — растение и лат. caedo — убиваю) — образуемые растениями, биологически активные вещества, убивающие или приостанавливающие размножение других организмов, главным образом — микробов. Обычно выделяются растениями в газообразном виде, к примеру, аллицин у лука и чеснока. Наличие фитонцидов играет крайне важную роль в формировании устойчивости растения к грибным заболеваниям.
Фитонциды имеют медицинское значение, из них изготавливаются некоторые препараты. За лето гектар лиственного леса выделят 2 кг фитонцидов, хвойного — 5 кг, можжевельника — 30 кг! Санатории часто располагаются в сосновых борах, где наблюдается повышенная концентрация фитонцидов. Вдыхание такого воздуха очень полезно при заболеваниях дыхательной системы инфекционной природы (когда возбудителями являются бактерии, грибы).

Значение голосеменных
Трудно переоценить значение голосеменных для человека, они очень важны. Голосеменные — источники высококачественной древесины, продуктов ее переработки. Являются звеном в цепи питания (продуцентами), основой многих биоценозов. Хвойные растения в больших количествах выделяют фитонциды, имеющие медицинское значение. Из смолы хвойных получают канифоль, скипидар, лаки. Кедровых орехи — это семена нескольких видов растений из рода сосна, которые употребляют в пищу.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Отдел Голосеменные

Голосеменные растения – группа высших семенных растений, у которых семязачатки образуются не внутри завязи пестика, как у цветковых растений, а в шишках, на поверхности чешуй.
Впервые выражение «голосеменные» использовал ботаник Бекетов. А.Н.
Термин «голосеменные» используется для отделения четырех нецветковых, семенных растений от класса «покрытосеменных».
Выделяют 4 отдела:
- Гинкговидные

- Гнетовидные

- Саговниковидные

- Хвойные

Общие особенности.
1.В отдел входят только древесные формы растений (деревья и кустарники).
2.Большинство являются вечнозелеными растениями (адаптации, повышающие зимостойкость).
3.Не имеют цветков и плодов, семена расположены открыто – в чешуях или шишках (мужские и женские).
4.В древесине отсутствуют настоящие сосуды (адаптация – листья игольчатой формы, для меньшего испарения влаги).
Систематика.

Общее строение.

Строение женской шишки сосны.
Весной на верхушках молодых побегов можно увидеть маленькие красноватые шишки. Это женские шишки. Женская шишка состоит из оси, или стержень, на котором располагаются чешуи. На чешуях женских шишек ничем не защищённые, как голые (отсюда название – голосеменные), лежат семязачатки, в каждом из них образуется яйцеклетка.
Строение мужской шишки сосны.
На тех же ветках, на которых расположены женские, находятся и мужские шишки. Они располагаются не на верхушке молодого побега, а у их основания. Мужские шишки мелкие, овальные, жёлтые и собраны в тесные группы.

Каждая мужская шишка состоит из оси, на которой также расположены чешуйки. На нижней стороне каждой чешуйке по два пыльцевых мешочка, в которых созревает пыльца – совокупность пылинок, в которых позже образуются мужские половые клетки – спермии.
Строение зрелой шишки сосны.
Оплодотворение у сосны происходит через год после попадания пыльцы на женские шишки. А высыпаются семена ещё через полгода, в конце зимы. К этому времени зрелая женская шишка становится бурого цвета и достигает 4-6 см.
Когда чешуи зрелой женской шишки раздвигаются, становится видно, что семена лежат попарно на верхней стороне чешуй, у их основания. Семена лежат, открыто, голо. Каждое семя сосны снабжено прозрачным пленчатым крылышком, обеспечивающее перенесение его ветром.
Процесс опыления и оплодотворения у сосны. (Цикл развития)
Размножение: половое – семенами.
Размножение осуществляется в два этапа: процесс опыления и процесс оплодотворения.
Процесс опыления
· Пыльца оседает на семязачатки женской шишки.
· Пыльца проникает внутрь семязачатка через пыльцевход.
· Чешуи смыкаются и склеиваются смолой.
· Подготовка к оплодотворению
· Пыльца, прорастая, образует спермии и пыльцевую трубку.
Процесс оплодотворения
Оплодотворение происходит в семязачатке через 12 месяцев после опыления.
· Спермий сливается с яйцеклеткой, в результате чего образуется зигота.
· Из зиготы развивается зародыш.
Шишка растёт и постепенно одревесневает, цвет её становится бурым. Следующей зимой шишки раскрываются и семена высыпаются. Они могут долго находиться в состоянии покоя и прорастают только в благоприятных условиях.
Очень своеобразно выглядят проростки сосны, когда они только что появились из семени. Это маленькие растеньица, у которых стебелек короче спички и не толще обыкновенной швейной иглы. На верхушке стебелька — пучок лучеобразно расходящихся во все стороны очень тонких иголочек-семядолей. Их у сосны не одна и не две, как у цветковых растений, а гораздо больше — от 4 до 7.
Таким образом, растения, относящиеся к отделу голосеменных, отличаются от всех остальных растений тем, что производят семена. Внутреннее оплодотворение, развитие зародыша внутри семязачатка и появление семени – главные биологические преимущества семенных растений, давшие им возможность приспособиться к наземным условиям и достигнуть более высокого развития, чем бессемянные высшие растения.

Значение в природе и в жизни человека.
Голосеменные составляют древесный ярус лесов умеренной зоны Они являются поставщиками кислорода и органических веществ, средой обитания животных и их пищей, хорошо очищают воздух не только от грязи, но е и от вредных микроорганизмов, участвующих в круговороте воды и других вен.
Из сердцевины саговника поникшего изготавливают крупу саго Крупные семена сибирского кедра, так называемые \"кедровые орешки\", употребляют в пищу.
Породы деревьев используются как топливо, строительные материалы (вагоностроение, кораблестроение, авиация, изготовление музыкальных инструментов и т.п.).
Сосновые почки, пихтовое масло, хвоя применяются для изготовления лекарственных препаратов.
В химической промышленности с голосеменных получают смолы и эфирные масла
Голосеменные являются декоративными и полезащитные растениями. в химической промышленности с голосеменных получают смолы и эфирные масла
Голосеменные
Голосеменные, или Acrogymnospermae — группа родственных растений, имеющих общих предков, схожих в характере размножения семенами, строении семязачатка, мужского и женского гаметофитов, опыления и оплодотворения. Именно семена отличают Голосеменные растения от папоротников и других споровых.
- большая часть растений, описываемых в этой статье — хвойные деревья и кустарники, родственники новогодней ёлки;
- все Голосеменные имеют два типа шишек: пыльцевые и семенные;
- до появления в результате эволюции семян животные питались только листьями, стеблями и корнями, которые содержат гораздо меньше питательных веществ;
- описанные ниже растения называются Голосеменными, но последние исследования показали, что они не являются единой группой;
- семенные растения хоть и размножаются семенами, но тоже образуют споры, и их жизненный цикл также состоит из спорофита (диплоидного) и гаметофита (гаплоидного).
Признаки голосеменных
Голосеменные включают в себя четыре из пяти линий семенных растений. Пятая линия — это Покрытосеменные, или Цветковые, их зародыш имеет замкнутое вместилище (околоплодник).
| Пять групп современных семенных растений | |||
| Отдел | Примеры | Ключевые характеристики | Примерное количество видов |
| Хвойные | Включают ели, сосны, пихты, тисы и др. | Гетероспоровые семенные растения с неподвижными сперматозоидами (спермиями), продвигающимися к яйцеклетке при помощи пыльцевой трубки. Листья игольчатые или чешуйчатые. Деревья, кустарники, кустарнички. Средняя продолжительность жизни около 68 лет. Семена производятся в шишках. | 630 |
| Цикадовые | Цикады | Разноспоровые растения с подвижными сперматозоидами, заключёнными в пыльцевую трубку, с помощью которой они подводятся к яйцеклетке. Листья перистые, как у пальм или папоротников. Вторичный рост стебля слабо выражен по сравнению с хвойными. Семена в шишках. | 306 |
| Гнетовые | Вильвичия удивительная | Разноспоровые растения с неподвижными сперматозоидами, подводимыми к яйцеклетке при помощи пыльцевой трубки. Единственное из голосеменных имеет сосуды. Деревья, кустарники, лианы, особая жизненная форма у вельвичии. Содержит 3 очень разные рода: эфедра, гнетум, вельвичия. | 65 |
| Гинкговые | Гинкго | Разноспоровые, сперматозоиды со жгутиками, но подводятся к яйцеклетке при помощи пыльцевой трубки. Листопадное дерево с веерообразными листьями и дихотомическим жилкованием. Семена напоминают сливы, но с дурно пахнущей оболочкой. Один род. | 1 |
| Ахрофита | Цветковые растения | Разноспоровые растения с неподвижными сперматозоидами, продвигающимися при помощи пыльцевой трубки. Семена заключены в плод, листья очень разнообразны по размеру и форме. Травы, лианы, кустарники, деревья. Около 14 тыс. родов. | Более 300 тысяч |
У Голосеменных яйцеклетки и зародыши (семена) лежат открыто:
- на семенной чешуе (видоизменённом побеге или спороносном листе — спорофилле) у хвойных;
- на мегаспорофиллах у многих других Голосеменных;
- единично — у тиса и гинкго.
Ткани спорофита покрывают семена не полностью. Семенные чешуи и мегаспорофиллы Голосеменных собраны в стробилы (видоизменённые побеги), называемые шишками . Ни цветков, ни плодов у них нет.

Жизненные формы голосеменных
Голосеменные — исключительно наземные растения. В подавляющем большинстве это крупные деревья, реже кустарники или кустарнички, едва превышающие по размеру мхи. Исключение составляют цикадовые — многолетние растения с толстым колоннообразным и почти всегда неветвящимся стволом, а иногда и клубневидным, частично погружённым в почву и напоминающим стебли папоротников.
Среди цикадовых есть эпифиты и очень мелкие растения, не превышающие 25 см, кустарнички и лианы, а жизненную форму вельвичии удивительной (Welwitschia mirabilis) трактуют как «взрослый проросток». Это своеобразное растение не имеет аналогов жизненной формы среди флоры нашей планеты. Есть среди Голосеменных и одно паразитическое растение из семейства подокарповые — паразитаксус опалённый (Parasitaxus ustus).

Голосеменные содержат и паразитическое растение: паразитаксус опалённый (Parasitaxus ustus). Автор: Tentaculata, CC BY 4.0
Самый крупный отдел Голосеменных – это Phylum, Coniferophyta, или хвойные растения, к которым относятся сосны, ели, пихты, кедры, тисы, лиственницы, кипарисы и др. Почти 40% мировых лесов представлены тайгой — лесом из хвойных пород.
Секвойядендрон гигантский, или мамонтово дерево (Sequoia sempervirens) — единственный современный вид рода Секвойя — хвойное дерево, произрастающее на северо-западе Калифорнии и юго-западе штата Орегон. Это самое высокое сосудистое растение, единичные экземпляры которого могут достигать 100 м. высоты.

Голосеменные растения — самое высокое дерево на Земле
Другое хвойное — сосна остистая межгорная (Pinus longaeva) — дерево с корой белого цвета. В горах Калифорнии это самое долгоживущее растение, одному из экземпляров которого 4900 лет. Хвойные встречаются в умеренном климатическом поясе. Они являются источником древесины, сырья для биотоплива, изготовления бумаги, добывания смолы, паклитаксела (используется для лечения рака) и других экономически важных продуктов.

Голосеменные — сосна остистая
Размножение Голосеменных
Голосеменные различаются по типу размножения и по способу роста. Например, саговники и гинкго имеют подвижные сперматозоиды, тогда как у хвойных и гнетовых сперматозоиды неподвижны — спермии . Все сперматозоиды переносятся к архегонию при помощи пыльцевой трубки, образующейся при прорастании пыльцевого зерна . Такой тип оплодотворения называется внутренним .
В жизненном цикле Голосеменных доминирует бесполая стадия — спорофит, а их гаметофит редуцирован ещё больше, чем у папоротников и мхов, и развивается внутри спорофита. Все Голосеменные разноспоровые, и их размножение не зависит от наличия воды. При этом их мега- и микроспоры часто бывают одинаковыми по размеру, а иногда микроспоры даже крупнее.

Развитие Голосеменных
В мегаспорангии ( нуцеллусе ) чаще всего образуется только одна клетка мегаспоры (материнская клетка зародышевого мешка), из которой в результате мейоза получается 4 мегаспоры. Обычно только одна мегаспора развивается в женский гаметофит ( зародышевый мешок ). Женский гаметофит образует яйцеклетку. В отличие от большинства Папоротникообразных, у семенных растений яйцеклетка не покидает мегаспорангий, и молодое растение развивается за счёт материнского спорофита.
Мегаспорангий семенных растений окружён стерильной оболочкой — интегументом , которого нет у Папоротникообразных. Интегумент, нуцеллус и зародышевый мешок с яйцеклеткой в совокупности называются семязачатком .

В микроспорангии образуются многочисленные материнские клетки микроспор (материнские клетки пыльцевых зёрен), из которых после мейоза возникает множество микроспор. Мужской гаметофит (пыльцевое зерно) развивается внутри оболочки пыльцевого зерна и образует имеющие жгутики сперматозоиды или безжгутиковые спермии (по сути подвижные ядра). В процессе опыления мужской гаметофит переносится к семязачатку, где он прорастает, образуя пыльцевую трубку, которая, удлиняясь, доставляет к яйцеклетке или подвижные сперматозоиды, или безжгутиковые неподвижные спермии.
В результате оплодотворения яйцеклетки сперматозоидом
или спермием получается зигота, развивающаяся в зародыш. После оплодотворения семязачаток развивается в семя. Семя Голосеменных состоит из:
- семенной кожуры , развивающейся из интегумента,
- многоклеточного зародыша,
- и (почти всегда) питающей ткани (чаще всего эндосперма).
Семя заменило одноклеточную спору папоротникообразных как средство распространения. Распространение семян Голосеменных растений осуществляется при помощи ветра и животных.
Кроме размножения семенами у голосеменных встречается и вегетативное размножение отводками.
Вывод. Характерные отличия размножения Голосеменных от размножения споровых растений:
- наличие семязачатка, в котором мегаспорангий (нуцеллус) окружён покровом (интегументом), формирующим пыльцевход (микропиле);
- развитие мужского и женского гаметофитов происходит не только внутри микро- и мегаспорагниев, но даже внутри спор;
- редукция антеридиев, а у некоторых Голосеменных (гнетовых) и архегониев;
- образование пыльцевой трубки для перемещения спермиев к яйцеклетке, что делает половой процесс независимым от наличия воды;
- женский заросток представляет собой эндосперм;
- развитие зародыша спорофита обычно происходит полностью на материнском растении;
- семя состоит из диплоидного зародыша (зачатка нового спорофита), покрова семени (производного спорофита предыдущего поколения) и эндосперма (женского заростка), который выполняет роль питательной ткани для развивающегося из зародыша проростка.
Строение Голосеменных
Спорофиты Голосеменных состоят из истинных органов: корня и побега со стеблем и листьями.
У них хорошо развита стержневая корневая система , в которой всегда есть главный и боковые корни, у более примитивных развиты и придаточные корни. Внутреннее строение корня сложное, он способен ко вторичному утолщению.

Органы Голосеменных на примере ели
Стебель Голосеменных с плохо развитой сердцевиной и мощными древесиной и корой характеризуется наличием сосудистых тканей, не организованных в проводящие пучки. В этом состоит отличие Голосеменных от Покрытосеменных. Сосуды ксилемы развиты только у гнетовых. Наличие камбия характеризует способность Голосеменных ко вторичному утолщению.

Листья современных Голосеменных весьма различны. При этом больше всего различаются листья самых примитивных современных Голосеменных (цикадовых и гинкговых). У цикадовых очень крупные, жёсткие перистые листья, напоминающие листья некоторых папоротников. У единственного представителя гинкговых — гинкго двулопастного (Ginkgo biloba), нижние листья веерообразны и имеют дихотомическое жилкование, как у большинства папоротников.
У большинства ныне живущих хвойных листья чешуевидные или игольчатые. Больший разброс в признаках листа имеется у гнетовых. Есть растения с редуцированными листьями (эфедра), с почти не отличающимися от листьев цветковых с перистым жилкованием (гнетум) и, наконец, с колоссальными ремневидными, растущими на протяжении всей жизни листьями (вельвичия).

У большинства голосеменных листья вечнозелёные, исключение — гинкго и некоторые хвойные (лиственница).
Классификация семенных растений
Старая классификация 1800 года объединяла все семенные растения в одно подразделение — сперматофоры (Spermatophyta) с двумя отделами: Голосеменные и Покрытосеменные. Голосемменные — это те растения, голые яйцеклетки которых расположены на плоских спорофиллах (например, сосновых шишках). Покрытосеменные — это цветущие растения с плодоножками-спорофиллами, образующими трубчатую замкнутую структуру. То есть их плоды — это зрелые плодоножки.
Эта классификация подчёркивает тесную связь всех семенных растений и указывает на происхождение Покрытосеменных от какой-то группы Голосеменных. Но если Покрытосеменные развились из Голосеменных, то Голосеменные не являются естественной группой, так как остаётся нерешённым вопрос о непосредственных предках Покрытосеменных. В кладистической терминологии неполная группа называется парафилетической. Следовательно, от понятия «Голосеменные» нужно отказаться.
Сегодня чаще используется другое деление живых семенных растений, без термина Голосеменные, на группы-отделы:
- Саговниковидные (Cycadophyta);
- Хвойные (Pinоphyta, или Cycadophyta);
- Гнетовидные (Gnetophyta);
- Гинкговидные (Ginkgophyta);
- Цветковые (Magnoliophyta).
Кроме этих групп существуют вымершие, наибольшее значение для понимания эволюции семенных имеют прогимноспермы и семенные папоротники . О них мы и поговорим подробнее.
Эволюция семенных растений
Жизненный цикл всех ранее рассмотренных сосудистых растений представляет собой чередование независимых гетероморфных поколений (полового — гаметофита и бесполого — спорофита). Недостаток его в том, что новый спорофит, развиваясь из зиготы, временно зависит от крошечного гаметофита с малым количеством питательных веществ. Так как гены гаметофита и эмбриона, а значит и нового спорофита, одинаковы, любая мутация может привести к их гибели. Так и происходит, большая доля молодых спорофитов не выживает.
Более выгодна стратегия семенных. Они способны использовать фотосинтетическую и абсорбционную способность предыдущего спорофита. Для того, чтобы это произошло, мегагаметофит должен быть сохранён внутри материнского спорофита. Это достигается путём сохранения мегаспоры внутри спорангия, где из неё и развивается гаметофит.

Соотношение гаметофитов и спорофитов растений
Такое положение мегагаметофита требует изменения и в микрогаметофите. Положение в стробилах на растении не позволяет использовать подвижные сперматозоиды для оплодотворения мегагамет, расположенных тоже в стробилах. Это препятствие было преодолено пространственным расположением — в вертикальных конусообразных спороносных побегах (стробилах, или шишках), позволяющих микроспорам переноситься ветром и опускаться на конус мегаспорангия.
Там микроспоры прорастают в микрогаметофит, производят антеридии и сперматозоиды, а также осуществляют оплодотворение. Само оплодотворение изменилось мало, причём влажный спорофилл заменил влажную почву. Эти изменения и породили первые семенные растения — семенные папоротники (Pteridospermophyta), эволюционировавшие в первые хвойные, ныне вымершие растения порядка Cordaitales. Этот метод оплодотворения существует у потомков этих вымерших групп растений — у саговников и гинкго. Они отличаются от других голосеменных наличием подвижных сперматозоидов.

В других линиях, ведущих к хвойным и цветковым растениям, мужские половые клетки стали неподвижными (спермии) и переносились к яйцеклетке путём прорастания микрогаметофита в пыльцевую трубку.
Эволюции семян предшествовала эволюция сосудистого камбия и сосудистых тканей. В некоторых случаях камбий возникал самостоятельно (древние плауновидные и хвощевидные), клетки в нём могли подвергаться радиальному продольному делению, что позволяло ткани камбия нарастать в окружности по мере накопления древесины. В отличие от хвощей и плаунов, новый тип камбия смог и до сих пор приводит к массивности деревьев, таких как гикори, секвойя, дуб. Видимо, этот камбий возник только один раз, в группе растений, которая дала начало всем древесным породам.
Только после этого появились семена и семенные растения — сперматофоры. Но до появления семян существовали и деревья, размножающиеся спорами, такие как папоротники. Учёные всё ещё не до конца уверены в эволюционных связях всех групп сохранившихся семенных растений и их вымерших предков. Одна из проблем в том, что из ископаемых растений невозможно извлечь ДНК.
Все голосеменные можно разделить на две группы:
- одна производит небольшое количество очень мягкой губчатой паренхимной древесины (моноксильная древесина). Имеет крупные сложные листья и радиально симметричные семена. Эти характеристики встречаются у саговниковых и их родственников;
- вторая характеризуется наличием твёрдой крепкой древесины с небольшой паренхимой (пикноксильная древесина), мелкими простыми листьями и приплюснутыми семенами (двунаправлено симметричными). Это характерно для хвойных и их родственников.
Видимо, хвойные развились из цикадовых, так как они очень похожи.
Прогимноспермные (Progymnospermophyta)
Важной группой растений, развившихся из риниофитов, были тримерофиты . Одна тенденция эволюции, начатая ими, представляла собой псевдомоноподиальное ветвление и первые шаги в эволюции мегафиллов (листьев с множеством разветвлённых жилок). Эта тенденция была продолжена в хвощах и папоротниках, которые развились из тримерофитов. Третья группа, развившаяся из тримерофитов, — это вымершие ныне прогимноспермные растения, названные так потому, что из них развились хвойные, саговники и некоторые другие Голосеменные.
Так же как у папоротников и хвощей, у прогимноспермных развивались мегафилльные листья. Однако значимой была и другая их особенность — эволюция сосудистого камбия с неограниченным потенциалом роста и способностью производить как вторичную ксилему, так и флоэму. Ещё 360 млн. лет назад сосудистый камбий, эволюционировавший в группе прогимноспермных, был способен претерпевать радиальные изменения, продольное деление и функционировать бесконечно долго.

Древний лес был похож на современную тайгу из хвойных пород деревьев. Они содержали в стволах трахеиды удлинённой формы, чаще с окаймлением из округлых ямок. Осевая паренхима же была слабой или отсутствовала. Лучи состояли из лежачих трахеид. Такая древесина должна была быть сильной, способной поддерживать большую массу и эффективно проводить воду и минералы.
Некоторые виды, такие как Трилобоксилон и Протеокалон, имели и пробковый камбий, производивший кору. Прогимноспермы были настоящими деревьями. Их стволы достигали 1,5 м. в диаметре и 12 м. в высоту.
Хотя древесина прогимноспермных походила на древесину хвойных, эти две группы вовсе не одинаковы. У первых не было семян и даже предшественников яйцеклеток. Виды, возникшие позже, были гетероспоровыми, но мегаспоры не удерживались в неразличимом ещё спорангии, как это происходит у семенных растений. Хотя листья и древесина прогимноспермных были довольно развиты, их размножение ещё было простым.
Порядок Аневрофитовидные, или Протоптеридиевидные (Aneurophytales, или Protopteridiales)
Содержит более реликтовые прогимноспермные, такие как Aneurophyton, Protopteridium, Proteokalon, Тетраксилоптерис, Трилобоксилон и Эосперматоптерис. Среди них были невысокие кустарники (Протоптеридий, Tetraxylopteris) и крупные деревья до 12 м. высотой.

Aneurophyton germanicum
Автор: Гедогедо, CC BY-SA 3.0
Все они имели сосудистый камбий, были способны ко вторичному росту, но их первичная ксилема составляла протостелу, как у риниофитов и тримерофитов. Аневрофиты ещё больше напоминали тримерофиты, между их ветвями были тонкие нити, которые нельзя было назвать листьями.
Порядок Археоптерисовидные (Archaeopteridales)
Это более «продвинутые» прогимноспермные, деревья высотой до 8,4 м. с обильной древесиной и вторичной флоэмой. Стебли Археоптериса имели сифоностелу, сердцевину, окружённую пучком первичной ксилемы, очень похожую на таковую у современных хвойных и двудольных покрытосеменных. Листья их внешне напоминали вайи папоротников, но на самом деле были системой ветвей со спирально расположенными простыми листьями. У A. macilenta жилкование их было лишь частичным, а у A. halliana полным, и их уже можно считать полноценными мегафиллами.

Археоптерис, реконструкция. Автор: Retallack
CC BY-SA 4.0
Размножение у археоптеридов было гетероспоровым. Мегаспоры диаметром до 300 мм и микроспоры диаметром в 30 мм, каждый тип спор производился в своих спорангиях — микро- и макроспорангиях. Спорангии были терминальными на коротких смешанных (стерильных и фертильных) ветвях, похожих на листья. Мегаспоры не удерживались внутри спорангия и семена не производились.
Эволюция семени
Исследование жизненного цикла вымерших растений на основе ископаемого материала трудно, но не невозможно. Если окаменевшие споры находились в свободном состоянии, то их идентифицировали как спорофиты, если же споры оказывались внутри спорангия, прикреплённого к листьям или древесине, то это уже определялось как предшественники гаметофита. К сожалению, споры не могут быть идентефицированы с гаметофитом (за исключением тех случаев, когда гаметофит микроскопичен и развивается внутри стенок спорангия).
В настоящее время самым ранним известным видом прогимноспермных растений с гетероспорией является Chauleria из среднего девона (примерно 390 лет назад). Окаменелый спорангий был обнаружен в одной из окаменелостей верхнего девона у Archaeosperma arnoldii. У него одна материнская клетка мегаспора дала начало одной большой жизнеспособной мегаспоре и трём маленьким. Этот мегаспорангий был окружён слоем покровной ткани, имелось большое микропиле (отверстие в покрове), оно позволяло сперматозоидам подбираться к яйцеклетке после того, как мегаспора превращалась в мегагаметофит и производила яйцеклетку. Этот процесс был похож на таковой у Покрытосеменных растений.
У Genomosperma kidstoni мегаспорангий был окружён стерильной тканью. Эта ткань была важна при улавливании путешествующих при помощи ветра микроспор. Вначале они приземлялись на любой части мегаспорангия, позже появилась «посадочная площадка» в верхней части мегаспорангия, она же была местом хранения или пыльцевой камерой. По мере того, как мегаспорангии эволюционировали в яйцеклетки со стерильным эпидермальным покрытием, другие талломы на соседних ветках стали видоизменяться в чашечки, которые позже стали плодоножками Покрытосеменных.
Одновременно микроспоры превращались в пыльцевые зёрна. Изменения происходили в характере их стенок, в способе прорастания, в природе производимого ими микрогаметофита. Сразу после верхнего девона, приблизительно 340 млн. лет назад стали обычным явлением четыре типа пыльцы. Некоторые из них сохранились в ископаемом виде настолько хорошо, что можно было увидеть, что эти зёрна уже имели внутреннее строение, удивительно похожее на современное у Голосеменных растений.
Подраздел птеридоспермы, или семенные папоротники (Pteridospermophyta)
Прогимноспермы дали начало не только хвойным, но и другой группе растений — цикадофитам. Они подразделяются на три группы:
- Pteridospermophyta (семенные папоротники, все вымершие),
- Cycadophyta (саговники, сохранившиеся),
- Cycadeoidophyta (цикадеоиды, все вымершие).
Самые ранние семенные папоротники появились в верхнем девоне, другие их группы возникли несколько позже. Не все они тесно связаны между собой, так как не являются потомками общего предка, поэтому не представляют собой таксона, а рассматриваются как отдельный уровень эволюции растений.

Голосеменные папоротники.
Автор: Retallack, CC BY-SA 3.0
Все семенные папоротники были деревьями с папоротниковидными листьями, которые несли семязачатки вместо спорангиев. Семязачатки были покрыты купулой (желеобразный колпачок), образованной из самого листа. Под купулой находился слой камер, сросшихся между собой боковыми краями, он получил название интегумента. В верхней части семязачатка находилось отверстие — микропиле. Под интегументом находился мегаспорангий, или нуцеллус, в котором развивалась одна крупная мегаспора. Было пространство, которое назвали пыльцевой камерой, в него попадали микроспоры и прорастали под защитой семязачатка.
Сперматозоиды папоротников сильно напоминали сперматозоиды цикадовых. Мегаспора семенных папоротников прорастала внутри мегаспорангия-нуцеллуса, образуя женский гаметофит с несколькими архегониями. Здесь же под защитой семязачатка совершалось оплодотворение яйцеклетки, а дальнейшее развитие спорофита проходило, вероятно, уже на земле и без периода покоя, характерного для семени семенных растений. Поэтому они считаются не семенными, а семязачатковыми растениями, так как настоящего семени они ещё не образовывали.

Многие из них внешне напоминали современные древовидные папоротники, за исключением того, что у них была древесина. Считается, что птеридоспермы произошли от Аневрофитов, потому что самые первые семенные папоротники, такие как Стеномиелон, имели протостелу из трёх рёбер. У более поздних видов Стеномиелона, Calamopitys и Lyginopteris, большая часть центральных клеток ствола становится паренхимой, а не трахеидосодержащей. У них было трёхцветное кольцо из сосудистых пучков, окружающих древесину. Такое положение встречается в стеблях всех Голосеменных и Покрытосеменных растений.
У папоротников был долгоживущий сосудистый камбий, производивший как ксилему, так и флоэму, а это тоже признак современных Голосеменных и Покрытосеменных. Хотя их древесина была похожа на древесину их предков (прогимноспермных), имелись и интересные отличия. Их трахеиды были намного длиннее и шире, на каждой из них находилось несколько рядов окаймляющих ямок, у хвойных трахеиды настолько узки, что на них может поместиться не более двух рядов ямок.
Древесинные лучи птеридосперма шириной во много клеток с большими клиньями паренхимы, поэтому древесина семенных папоротников была более мягкая и менее плотная, чем древесина хвойных и прогимноспермных. Такая древесина называется моноксильной, она встречается у саговников и цикадовых.
Стебли семенных папоротников имели толстую кору с радиальными пластинками склеренхимы, расположенными чуть ниже эпидермиса. Внутренняя кора содержала секреторные каналы. У более старых деревьев пробковый камбий и кора формировали наружную часть вторичной флоэмы. И вторичная и наружная кора сбрасывались растением совместно.
Хвойные (Pinоphyta, или Cycadophyta)
Поскольку хвойные растения для нас привычнее, чем другие Голосеменные, рассмотрим их в первую очередь. Они разнообразны (примерно 50 родов и 550 видов), все являются деревьями или кустарниками от умеренных до гигантских размеров. Среди хвойных нет лиан (есть стланики), трав или однолетних растений. У них не бывает луковиц или корневищ.

Листья хвойных — это простые иглы или чешуйки. Листья большинства хвойных многолетние, у агафиса и араукарии они остаются даже на очень старых растениях. Жилкование листьев хвойных часто бывает простым — с одной или двумя жилками, у чешуевидных листьев это несколько параллельно расположенных жилок. Жилки имеют эндодерму и ткань, называемую трансфузионной, состоящей из клеток трансфузионной паренхимы и более или менее кубовидных трахеид. Трахеиды содержат округлые выступающие ямки.
Трансфузионная паренхима смешивается с трахеидами и образует трёхмерный рисунок из двух типов клеток, облегчающий перенос веществ между обычными сосудистыми тканями и мезофиллом вне эндодермы.Как и у их прародителя Архиоптериса, у современных хвойных отсутствуют сосуды, а флоэма состоит из ситовидных клеток.

Голосеменные — хвойные
У всех хвойных деревьев есть пыльцевые и семенные шишки, у большинства видов они одревесневают, а у можжевельника и подокарпа они внешне напоминают плоды цветковых растений. Хвойные деревья — успешная группа растений, образующая обширные леса, охватывающие площадь более 17 млн. км². Большую площадь они занимают в Северном полушарии.
В качестве образца всегда принято рассматривать род Сосна. На сегодняшний день существует более 100 видов сосен, все их можно найти в США. Сосны и ели, которые принадлежат к одному семейству, образуют обширные хвойные леса в Северном полушарии, лежащие между арктическими пустынями, лиственными лесами умеренных широт и степями на юге. В недавнем прошлом сосны были рассажены в Южном полушарии, и там они тоже стали широко распространёнными.

Голосеменные, хвойные: тис
Морфология сосны
Как и большинство хвойных деревьев, сосны имеют два типа побегов, каждый из которых характеризуется наличием своих типов листьев. Крошечные бумажные листья появляются на длинных побегах, а в их пазухах располагаются короткие побеги, производящие знакомые нам игольчатые листья. Листья имеют много ксероморфных признаков:
- толстая кутикула;
- погружённые в ткань устьица;
- цилиндрическая узкая форма.
Жёсткие листья-иголки сосен растут группами от 2 до 5 штук. Из всех хвойных только сосны имеют сгруппированные листья. Их толстая кутикула и углублённые устьица — это приспособления к минимизации испарения воды. Это важно потому, что многие хвойные растут на слое вечной мерзлоты, что затрудняет корням доступ к воде.

Листья и другие части спорофита сосны имеют каналы, окружённые клетками, выделяющими смолу. Смола отпугивает насекомых и уменьшает грибковые атаки. Из смолы некоторых сосен делают скипидар и канифоль, используемый для струнных смычковых инструментов.
Древесина сосен считается мягкой из-за отсутствия некоторых жёстких типов клеток. Толстая кора сосен — это приспособление для выживания во время пожаров и минусовых температур. Некоторые шишки на самом деле не выпускают семена, если не подвергаются воздействию огня.

Туя. Какие Голосеменные!
Структура органов размножения сосен
Все семенные, как и споровые растения, производят два типа спор (микро- и макроспоры), которые дают начало двум типам гаметофитов. Мужские гаметофиты — микрогаметофиты (пыльцевые зерна) сосен развиваются из микроспор, которые появляются в результате подвергшейся мейозу материнской клетки. Мужские сосновые шишки обычно имеют длину от 1 до 4 см и состоят из мелких бумажных чешуек, расположенных по спирали или в виде завитков. Пара микроспорангиев формируются в виде мешочков внутри каждой чешуи. Многочисленные материнские микроспоровые клетки подвергаются мейозу, каждая из которых превращается в четыре микроспоры. Подвергаясь мейозу, многочисленные материнские микроспоровые клетки, производят по четыре пыльцевые зерна, каждое с парой воздушных мешочков, которые придают им дополнительную лёгкость при перенесении их ветром. Скопление мужских сосновых шишек на одном дереве может производить более миллиона пыльцевых зёрен.

Жизненный цикл Голоеменных на пртмере сосны
Женские сосновые шишки обычно образуются на верхушках побегов того же дерева, где появляются и мужские шишки. Они крупнее мужских, их чешуя при созревании семян одревесневает. В основании каждой чешуи развивается 2 яйцеклетки
Женский гаметофит в зрелом возрасте может состоять из тысяч клеток с двумя-шестью архегониями, образующимися на микропилярном конце. Каждый архегоний содержит яйцеклетку настолько большую, что её можно увидеть без микроскопа.

Представители Голосеменных: можжевельник
Оплодотворение и формирование семян сосны
Для созревания женских шишек сосны обычно требуется два или более сезонов. Поначалу они красноватого или пурпурного цвета, но вскоре становятся зелеными и коричневыми, после созревания в первую же весну чешуя раздвигается. Весной пыльцевые зерна, несомые ветром, дрейфуют вниз, некоторые попадают в липкую жидкость, сочащуюся из микропиле. Жидкость втягивает пыльцевые зёрна внутрь, к вершине нуклеуса, и вскоре после этого чешуя снова закрывается.

Строение семени Голосеменных
Во время роста пыльцевой трубки одна из четырех клеток пыльцевого зерна, генеративная клетка, делится путем митоза, причём одна из полученных двух клеток делится еще раз. Эти последние две клетки функционируют как сперматозоиды. Проросшее пыльцевое зерно с его двумя сперматозоидами является зрелым мужским гаметофитом, он гаплоиден и устроен намного проще, чем у папоротников.
Примерно через 15 месяцев после опыления пыльцевая трубка поступает в семяпочку через микропиле и выполняет его содержимое. Один сперматозоид соединяется с яйцеклеткой, образуя зиготу. Это даёт начало поколению спорофита. Другие сперматозоиды и клетки гаметофита пыльцевого зерна вырождаются. Зигота развивается в эмбрион, окружается женскими клетками гаметофита, которые обеспечивают его питание.
Кожный покров затвердевает, образуя семенную оболочку. Зрелый зародыш хвойных имеет такое же строение, как и у Покрытосеменных, у него есть корешок, гипокотель, эпикотель и семядоли. Но семядолей всегда много, а не одна или две, как у Покрытосеменных. Семя тоже напоминает семена Цветковых, но образуется оно не в плоде, а в конусе. У двух хвойных деревьев семена становятся мясистыми и ярко окрашенными — красными у подокарпа и синими у можжевельника.
После распространения и прорастания семени молодой спорофит следующего поколения превращается в дерево.
Саговники (Cycadophyta)
Это самая древняя группа Голосеменных растений. Современные саговники часто путают либо с папоротниками, либо с молодыми пальмами. У них толстые стволы и перистые сложные листья-ветви. Большинство саговников — это низкорослые растения, высотой менее 1 или 2 м., однако Микрозамия может достигать 18 м. высоты. Ствол покрыт корой и стойкими листовыми основаниями, которые остаются на растении даже после опадания листьев.
Внутреннее строение стебля саговника похоже на стебли семенного папоротника: толстая кора, содержащая секреторные протоки, окружает небольшой участок моноксильной (паренхимной) древесины. Их трахеиды длинные и широкие, а лучи массивные. Даже очень старые стебли имеют только небольшое количество древесины, а механическая поддержка растения обеспечивается в основном жёсткими основаниями листьев. Сердцевина саговников содержит секреторные каналы.
В отличие от семенных папоротников, листья саговников не несут яйцеклеток, вместо этого растения производят семенные и пыльцевые колбы на разных растениях — все саговники двудомны. Пыльцевые колбочки состоят из спирально расположенных щитовидных микроспорофиллов, несущих скопления микроспорагиев. При прорастании пыльцевые зёрна образуют разветвлённую пыльцевую трубку и крупные многожгутиковые сперматозоиды.

Cycas circinalis.
Автор: Jo Weber, общественное достояние
Семенные шишки изменчивы, самыми реликтовыми считаются шишки вида Cycas revoluta, у них семенной конус представляет собой рыхлое крупное скопление листовидных перисто-сложных мегаспорофиллов. От 6 до 8 крупных яйцеклеток находится у их основания, верхняя часть мегаспорофилла листовидная, похожая на органы, несущие яйцеклетки у семенных папоротников.
Более производными считаются мегаспорофиллы Zamia floridana. Они имеют форму щитков и несут по две яйцеклетки, а сами составляют довольно компактный конус. Яйцеклетки саговников похожи на яйцеклетки семенных папоротников, они заключены в васкуляризованный мегаспорангий с кожным покровом.

Голосеменные: саговник
В настоящее время Cycadophyta представлены девятью или десятью родами со 100 видами. Они растут в тёплом климате и не могут выдерживать морозных зим. Почти все они жители тропиков с небольшим ареалом: некоторые встречаются на Кубе, в Мексике, на юге США, другие — в Австралии, третьи — в Юго-Восточной Азии и Африке. Ранее они были распространены повсеместно, но с изменением климата во многих районах вымерли.
Отдел Гинкговидные (Ginkgophyta)
Содержит единственный сохранившийся до нашего времени вид — гинкго двулопастный (Ginkgo biloba). Может показаться необычным возводить целую дивизию для одного солдата, однако этот вид Голосеменных весьма необычен. Внешне он очень похож на двудольное дерево с толстым стволом и множеством веток. Но его древесина похожа на древесину хвойных, в ней отсутствуют сосуды и осевая паренхима. У гинкго есть широкие двулопастные вееровидные листья, но с реликтовым дихотомическим жилкованием, как у семенных папоротников. Как и у хвойных, у гинкго в наличии оба вида побега — короткие, несущие большую часть листьев, растущих группами (по 2-4) и длинные побеги с одиночно расположенными листьями. Гинкго — двудомное дерево, его семена расположены голо, но шишки у него отсутствуют. Вместо этого яйцеклетки появляются парами на концах короткого стебля.

Вот такие они Голосеменные!
Пыльца производится в органе, похожем на серёжки — колосовидные соцветия покрытосеменных, они имеют стебель и несколько спорангиофоров, каждый из которых содержит по 2 микроспорангия. Подобно яйцеклеткам птеридоспермов и саговников, яйцеклетки гинкго крупные (1,5-2,0 м в диаметре) и развивают трёхслойную семенную оболочку.
Гинкго двулопастный — очень популярное декоративное растение. Осенью его листья становятся ярко-жёлтыми. Разводить предпочитают мужские деревья, так как женские производят семена, наружный мясистый слой которых выделяет маслянистую кислоту с очень неприятным гнилостным запахом. Хотя семена дерева съедобны и неплохи на вкус, похожи на зелёный горох. Однако распознать пол дерева можно только на 25-30 год жизни, когда разовьются их генеративные органы.

Ginkgo biloba. Автор: Agnieszka Kwiecień, Nova, CC BY-SA 4.0
Точные предки гинкго неизвестны, но предположительно это была одна из групп семенных папоротников или их близкий родственник. Широко распространены гинкго были в мезозое, особенно в юрском периоде (примерно 170 млн. лет назад), окаменелые остатки его листьев и древесины можно найти практически во всех уголках земного шара, особенно в высоких широтах, таких как Аляска, Канада, Сибирь, а также в Патагонии, Южной Африке и Новой Зеландии. Тогда существовало не менее 15 родов этого отдела.
Гинкго начали вымирать в начале третичного периода, два его вида исчезли в эпоху плиоцена, примерно 10-12 млн. лет назад. Сохранившийся гинкго двулопастный в естественных условиях растёт редкими очагами в Китае и Японии.

Голосеменные: гинкго двулопастный
Отдел Гнетовидные (Gnetophyta)
Отдел содержит три порядка загадочных растений, в каждом из них по одному роду:
- Гнетовые (Gnetum) с 30 видами;
- Эфедровые, или Хвойниковые (Ephedrales) — около 40 видов;
- Вельвичия удивительная (Welwitschia mirabilis) — единственный вид рода Welwitschia.
Гнетовые — главным образом лианы, реже небольшие кустарники и деревья, растущие в тропических областях Земли — в Юго-Восточной Азии, тропической Африке и бассейне Амазонки. У них широкие листья, похожие на листья двудольных покрытосеменных растений.
Эфедровые — жёсткие кустарники и кустарнички, распространённые в пустынных районах Мексики, в юго-западной части США и в сухих горах Южной Америки. Их листья редуцированы до чешуек.
Немногочисленные представители рода вельвичия существуют только в пустынях Южной Африки или в культуре. У них короткий широкий стебель и только два листа. Но листья растут постоянно из прикорневой меристемы, становясь всё длиннее.

Голосеменные: вельвичия
Все три рода необычны тем, что представляют собой Голосеменные растения с сосудами в древесине. В эту группу их отнесли, потому что думали, что они связаны с примитивными Покрытосеменными, однако их сосудистые элементы развились из трахеид с окаймлением из округлых ямок, тогда как у Покрытосеменных сосуды появились из трахеид лестничного типа. Кроме того считается, что Покртосеменные эволюционировали от предков, лишённых сосудов, а сосуды развились после появления цветка.
В отличие от других Голосеменных, шишки гнетофитов сложны и содержат прицветники. Семенные шишки содержат дополнительные слои ткани вокруг яйцеклеток. Эта ткань интерпретируется по-разному, то как дополнительный покров, то как прицветник или спорофилл.
Их пыльца легко опознаваема, она имеет веретенообразную форму и узкие гребни. Её ископаемые останки датируются триасовым периодом. Но ни пыльца, ни другие окаменелости не помогли определить эволюционные связи этой группы растений. Есть версия, что гнетофиты и Покрытосеменные составляют две родственные группы с общим предком, но для подтверждения этой теории пока ещё слишком мало данных.
Значение Голосеменных
Голосеменные, как и все зелёные растения Земли, участвуют в процессе фотосинтеза, поглощают углекислый газ и обогащают воздух кислородом. Хвойные леса сохраняют влагу, препятствуя эрозии почв, и выделяют в воздух фитонциды (от греч. фитон — «растение» и лат. цидо — «убивают») — особые биологически активные вещества, уничтожающие микробов и препятствующие их размножению.
Значение Голосеменных растений в жизни и хозяйственной деятельности человека:
- использование древесины в строительстве и в качестве поделочного материала, как сырья для изготовления мебели, музыкальных инструментов;
- получение бумаги из древесины ели;
- производство искусственных тканей из древесины сосны (вискоза);
- получение спиртов, пластмасс, лаков и другого сырья для промышленности;
- изготовление пищевого масла из семян кедровой сосны и использование «кедровых орехов» в пищу;
- получение витаминов (витамин С, каротин);
- получение смолы и её производных: скипидара, канифоли;
- использование некоторых голосеменных растений (например, сосны) для закрепления почвы и предотвращения эрозии;
- высадка по периметру полей защитных лесополос, задерживающих таяние снегов (повышение влажности почвы) и препятствующих сильным ветрам, которые могут повредить посевы;
- озеленение дворов и улиц.
Хвойные — основные лесообразующие деревья России. А леса — это места обитания многих животных и растений. В тайге России наибольшие площади занимают леса из лиственницы, затем сосны и ели. Древесина лиственницы отличается особой прочностью и долговечностью, она устойчива к гниению.
https://studarium.ru/article/33
https://botanikagsk.ucoz.ru/index/otdel_golosemennye/0-10
https://tvoiklas.ru/golosemennie/